Že nekaj časa je aktualna problematika rasti števila prebivalstva, ki se bo do leta 2050 približalo desetim milijardam, zaradi česar bo treba več pozornosti nameniti pridelavi hrane. Poleg tega si v kmetijstvu prihodnosti želimo okolju prijazne, a hkrati učinkovite pridelave kakovostnih pridelkov. Zato na Oddelku za biotehnologijo in sistemsko biologijo Nacionalnega inštituta za biologijo iščemo rešitve za zmanjšanje izgub pridelka zaradi različnih okoljskih dejavnikov, med drugim tudi napada škodljivcev. Eden izmed načinov je preučevanje mehanizmov odpornosti v rastlinah. V naših raziskavah mehanizme odpornosti preučujemo pri krompirju, in to v interakciji z virusom Y krompirja, ki je eden izmed ekonomsko najpomembnejših rastlinskih virusov, saj okužba rastlin krompirja na polju s tem virusom lahko vodi v kar 80-odstotno izgubo pridelka (Slika 1). Mehanizme odpornosti lahko preučujemo na različne načine, ena izmed novejših, tako imenovanih prebojnih tehnologij, pa so genetski biosenzorji.

Slika 1: Okužba krompirja genotipa Rywal z virusom Y krompirja. Krompir je ena izmed najpomembnejših poljščin z letno proizvodnjo okoli 350 milijonov ton, zato je preučevanje interakcije med krompirjem in virusom Y krompirja ključnega pomena za zagotovitev učinkovitega in okolju prijaznega sistema zaščite rastlin. Okužba krompirja na polju s tem virusom močno prizadene pridelek in lahko vodi v kar 80-odstotno izgubo pridelka. Na sliki so rastline krompirja, ki smo jih v laboratoriju okužili z virusom Y krompirja. Okuženi listi porumenijo, se posušijo in nato odpadejo.
Kaj je genetski biosenzor in kako deluje?
Genetski biosenzor je orodje, ki zazna izbrano spremembo v celici in o tej spremembi poroča na način, da odda merljiv signal. Poznamo različne tipe biosenzorjev, eni izmed najpogosteje uporabljenih pa temeljijo na fluorescenci, ki jo odda fluorescenčni protein, ko zazna spremembo okolja v celici. Eden izmed takih, ki zaznava oksidirajoče okolje v celici, je na oksidirajoče okolje občutljivi zeleni fluorescenčni protein (angl. redox sensitve green fluorescent protein). Ko v celici nastanejo reaktivne kisikove spojine, se struktura tega fluorescenčnega proteina spremeni, zato se aktivira in zasveti. Da pa biosenzor lahko poroča o dogajanju v celici, ga moramo najprej vnesti v celice rastlin s postopkom transformacije. Ko se okolje v določeni celici spremeni v oksidirajoče zaradi nastanka kisikovih spojin, se biosenzor aktivira, kar opazimo kot fluorescenco (Slika 2a).
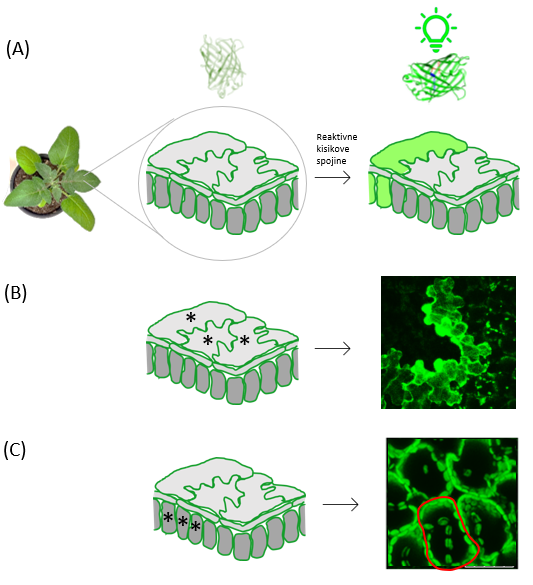
Slika 2: Na redoks stanje občutljivi biosenzor je občutljiv na oksidirajoče okolje v celici (nastanek reaktivnih kisikovih spojin). (A) Da biosenzor lahko poroča o dogajanju v celici, ga moramo najprej vnesti v celice rastlin. Ko pride do nastanka reaktivnih kisikovih spojin v določeni celici, se biosenzor aktivira, pri čemer spremeni svojo strukturo, in celica zasveti (označeno z zeleno). (B) Aktivacija na redoks stanje občutljivega biosenzorja v celicah povrhnjice, kot jo vidimo pod mikroskopom. Celice, v katerih je prišlo do aktivacije senzorja, so označene z zeleno. (C) Aktivacija na redoks stanje občutljivega biosenzorja v kloroplastih mezofilnih celic, kot jo vidimo pod mikroskopom. Kloroplasti, v katerih je prišlo do aktivacije senzorja, zasvetijo (označeno zeleno). Posamezna mezofilna celica je označena z rdečo.
V prečnem prerezu lista krompirja, kot je prikazan na Sliki 2, se na površini nahajajo celice povrhnjice, ki imajo obliko sestavljanke, medtem ko se globlje v tkivu nahajajo mezofilne celice (celice, v katerih se vrši fotosinteza), ki so stolpičaste strukture. Če pride do aktivacije biosenzorja v celici povrhnjice, to pod mikroskopom vidimo, kot je prikazano na Sliki 2b. Do aktivacije biosenzorja pa lahko pride tudi globlje v tkivu, v mezofilnih celicah in v tem primeru to s ptičje perspektive opazimo, kot je prikazano na Sliki 2c (posamezna celica je označena z rdečo). Zakaj s te perspektive? Ker je to zorni kot, s katerega opazujemo tkivo pod mikroskopom. V primeru na Sliki 2c je prikazana aktivacija biosenzorja v kloroplastih mezofilnih celic. Kloroplasti so razporejeni ob rob mezofilnih celic, zaradi česar nazorno označujejo posamezne celice oziroma meje med njimi. Biosenzor pa lahko vnesemo tudi v katere koli druge organele v celici, kot so na primer jedro, mitohondrij in endoplazmatski retikulum, in tako opazujemo dogajanje v točno določenem celičnem organelu.
Kaj je glavna prednost genetskih biosenzorjev?
Biosenzorji omogočajo spremljanje procesov v živo skozi prostor in čas in to celo na nivoju posamezne celice. To pomeni, da lahko z genetskimi biosenzorji spremljamo prenos signala iz celice, v kateri je prišlo do aktivacije biosenzorja, v sosednje celice (Slika 3). In ravno to je glavna prednost, ki biosenzorje loči od prejšnjih, starejših metod za spremljanje dogajanja v celicah. S starejšimi metodami smo namreč dogajanje v celicah spremljali na način, da smo tkivo odvzeli, nato pa iz tkiva izolirali izbrano molekulo. Na ta način smo procese v celici ustavili v trenutku, ko smo tkivo odvzeli. Tako smo procese lahko spremljali samo v točno določenem trenutku in v točno določenem tkivu. Medtem pa biosenzorji omogočajo spremljanje dogajanja na nivoju posamezne celice v realnem času in v vseh tkivih hkrati.
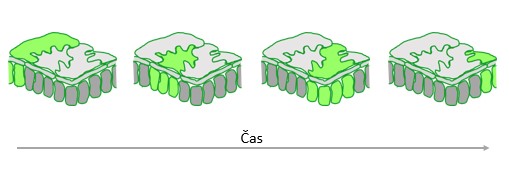
Slika 3: Biosenzorji omogočajo spremljanje dogajanja na nivoju posamezne celice v realnem času. Na sliki vidimo prenos signala iz celic, v katerih je prišlo do aktivacije biosenzorja, v sosednje celice. Ob določenem času aktivirane celice povrhnjice in mezofilne celice so označene z zeleno. Na sliki je prikazan prečni prerez lista, pri katerem se na površini nahajajo celice povrhnjice, ki imajo obliko sestavljanke, globlje v tkivu pa se nahajajo mezofilne celice, ki so stolpičaste strukture.
Kako smo se lotili preučevanja mehanizmov odpornosti?
Po tem, ko smo senzor vnesli v na virus odporne in občutljive rastline krompirja, smo rastline okužili z virusom Y krompirja in spremljali dogajanje v listih rastlin na nivoju posamezne celice. Dlje stran od mesta okužbe smo odkrili mezofilne celice, v katerih je prišlo do aktivacije biosenzorja, česar nismo opazili v na virus občutljivih rastlinah. Na spodnji sliki je prikazana aktivacija biosenzorja v kloroplastih rastlin mezofilnih celic, kot jo vidimo po računalniški analizi slike, ki smo jo posneli z mikroskopom. Pri analizi vsakemu kloroplastu dodelimo neko vrednost, ki priča o tem, kako močno je senzor aktiviran (kako močno sveti). To vrednost nato prikažemo na barvni skali. Rdeča barva priča o močni aktivaciji biosenzorja (senzor zasveti, ker je v kloroplastih prišlo do nastanka reaktivnih kisikovih spojin), modra barva pa nam pove, da v tem kloroplastu senzor ni aktiviran (senzor ne zasveti, ker v kloroplastu ni prišlo do nastanka reaktivnih kisikovih spojin). Glede na to, da smo mezofilne celice, v katerih je prišlo do aktivacije biosenzorja, opazili samo v na virus odpornih rastlinah, smo zaključili, da so te celice vključene v zaustavitev virusa, oziroma, da omogočajo odpornost krompirja na virusno okužbo. Bolj podrobni rezultati raziskave, ki predstavljajo pomemben tehnološki korak naprej za kmetijstvo prihodnosti, so dostopni v znanstveni reviji New phytologist.
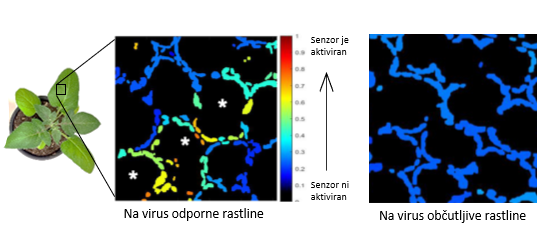
Slika 4: Aktivacija senzorja v kloroplastih rastlin mezofilnih celic. Senzor smo vnesli v na virus odporne rastline krompirja in v na virus občutljive rastline krompirja, jih okužili z virusom Y krompirja in spremljali dogajanje na nivoju posamezne celice. Na sliki je prikazana aktivacija biosenzorja v kloroplastih rastlin mezofilnih celic, kot jo vidimo po računalniški analizi slike, ki smo jo posneli z mikroskopom. Pri analizi vsakemu kloroplastu dodelimo neko vrednost, ki priča o tem, kako močno je senzor aktiviran, kar prikažemo na barvni skali. Rdeča barva: v tem kloroplastu je senzor aktiviran. Modra barva: v tem kloroplastu senzor ni aktiviran. Mezofilne celice, v katerih je prišlo do aktivacije senzorja v kloroplastih teh celic, so označene z asteriskom.
Niso pa to edini biosenzorji, ki smo jih razvili, v teku je še kar nekaj raziskav, v katerih razvijamo različne tipe senzorjev, s katerimi preučujemo različne tipe interakcij med rastlinami krompirja in različnimi okoljskimi dejavniki. Na primer, trenutno je v teku razvoj biosenzorjev treh glavnih rastlinskih hormonov (salicilna kislina, jasmonska kislina in etilen), ki so vključeni v odziv rastline na napad škodljivcev. Ti senzorji se aktivirajo ob prisotnosti omenjenih rastlinskih hormonov. Uporabljamo jih za preučevanje interakcije med rastlinami krompirja in virusom Y krompirja ter med rastlinami krompirja in koloradskim hroščem, ki poleg virusa Y krompirja povzroča znatno izgubo pridelka.
Kateri so glavni izzivi pri uporabi senzorskih rastlin?
Če želimo izkoristiti glavno lastnost biosenzorjev, ki je spremljanje procesov z veliko prostorsko in časovno ločljivostjo, moramo dogajanje opazovati v celih rastlinah. Zaradi pomanjkanja mikroskopov, ki bi omogočali spremljanje dogajanja v celih rastlinah, v praksi tega ni vedno enostavno doseči. Zato je v številnih laboratorijih po svetu, ki takih mikroskopov nimajo, treba pripraviti preparate rastlin, ki pa ne omogočajo spremljanja dogajanja v celicah v daljšem časovnem obdobju. Poleg tega smo v primeru priprave preparata omejeni na opazovanje le točno določenega območja tkiva. Zato v prihodnosti pričakujemo razvoj različnih tipov mikroskopov, ki bodo omogočali opazovanje celih rastlin, kar bo občutno povečalo tako časovno kot tudi prostorsko ločljivost.
Dodatno težavo predstavlja avtofluorescenca. Avtofluorescenca je fluorescenca nekaterih molekul, ki so naravno prisotne v tkivu. Za razliko od živalskih tkiv, pri katerih je avtofluorescenca manj pogosta, pa rastlinska tkiva vsebujejo več struktur, ki omogočajo sipanje in absorpcijo svetlobe in tako povzročajo avtofluorescenco. Rastlinsko tkivo vsebuje organske molekule, podobne tistim, ki jih najdemo v živalskem tkivu, poleg tega pa vsebuje še celulozo in lignin (sekundarna stena), klorofil (kloroplasti), antocianine in flavonole (vakuola), sporopolenin (zunanja stena pri cvetnem prahu), kutin (stena epidermalnih celic), škrob (amiloplasti) in ostale pigmente, ki absorbirajo svetlobo. Glavni vir avtofluorescence v rastlinah predstavljajo fenolne spojine ter klorofil v kloroplastih. Ker te molekule absorbirajo svetlobo v različnih delih spektra svetlobe, to predstavlja težavo pri ločevanju avtofluorescence od fluorescence različnih fluorescenčnih proteinov, ki prav tako fluorescirajo v enakih delih spektra svetlobe kot zgoraj omenjene molekule. Ker so fluorescenčni proteini glavna komponenta biosenzorjev, avtofluorescenca tako lahko moti signal biosenzorjev.
Tretja težava je priprava rastlin z vnesenimi senzorji, ki je za večino rastlinskih vrst dolgotrajen postopek, za določene rastlinske vrste pa je (trenutno) celo neizvedljiva. Kot že omenjeno v prejšnjih poglavjih, senzor v rastlino vnesemo s postopkom transformacije. Transformacija rastlin v večini primerov poteka preko vnosa senzorja z agrobakterijo, ki pa ni uspešna pri vseh rastlinskih vrstah. Še večjo težavo pa predstavlja vzgoja rastlin po transformaciji, ki ima za večino rastlinskih vrst nizek odstotek uspešnosti. Razlog za to je dolgotrajen in zahteven postopek transformacije in vzgoje rastlin po transformaciji, ki zahteva specializirano eksperimentalno znanje ter optimizacijo protokola za vsako rastlinsko vrsto posebej.
V katero smer gre razvoj biosenzorjev in kaj pričakujemo v prihodnosti?
Z biosenzorji lahko spremljamo že veliko različnih procesov v celici. Poleg reaktivnih kisikovih spojin trenutno lahko zaznavamo kalcij, ki je vključen v praktično vse življenjske procese, od rasti in razvoja pa do odziva na različne okoljske dejavnike. Poleg tega obstaja kar nekaj genetskih biosenzorjev za zaznavanje hormonov, ki so vključeni v razvoj in rast rastline, kot so citokinini, avksini, abscizinska kislina in giberelinska kislina. Ostaja pa še kar nekaj procesov, za katere je nabor senzorjev omejen in eden izmed njih je biotski stres. Trenutno gre razvoj v smeri priprave biosenzorjev za detekcijo treh glavnih rastlinskih hormonov, ki so vključeni v odziv rastline na škodljivce, in sicer salicilne kisline, jasmonske kisline in etilena, za katere še ni bilo razvitih veliko biosenzorjev.
Na polju so rastline ob istem času izpostavljene različnim tipom stresa, kot so na primer suša, vročina, slanost in napad škodljivcev. Kombinacije različnih tipov stresa imajo velik vpliv na rast rastlin in donos pridelka, kar postaja še bolj problematično zaradi podnebnih sprememb. Zato gre razvoj tudi v smeri razvoja kompleksnejših biosenzorjev, s katerimi bomo lahko zaznavali različne procese v celici hkrati in tako dobili boljši vpogled v odziv rastline na kombinacijo različnih tipov stresa. Uporaba različnih tipov biosenzorjev hkrati pa ne omogoča samo pridobivanja informacije o različnih spremembah v celici, temveč tudi spremljanje določene spremembe v različnih celičnih organelih. Komunikacija med organeli uravnava številne procese v celici, med drugim je vključena tudi v mehanizme odpornosti, zato je spremljanje sprememb v različnih celičnih organelih hkrati izjemnega pomena.
Biosenzorji postajajo vedno bolj občutljivi na najmanjše spremembe, hkrati pa dovolj robustni, da so uporabni kljub motečim komponentam v celici, zaradi česar bomo lahko zaznali že zelo majhne spremembe v celici, ki smo jih do sedaj spregledali. Pričakujemo pa tudi razvoj biosenzorskih poljščin, ki trenutno zaostaja za razvojem biosenzorskih modelnih rastlin (navadni repnjakovec in tobak), saj je vnos biosenzorjev v večino poljščin težaven in dolgotrajen. V teku je odkrivanje novih, bolj učinkovitih načinov za transformacijo in regeneracijo transformiranih senzorskih rastlin. Biosenzorske poljščine bodo omogočile preučevanje mehanizmov odpornosti v točno določeni poljščini, na primer krompirju, rižu ali pšenici in ne le v modelnih rastlinah, kar bo vodilo v razvoj odpornejših poljščin.