Britanski naravoslovec Alfred Russel Wallace je med svojim popotovanjem po jugovzhodni Aziji sredi 19. stoletja opazil, da obstaja med otoki današnje Indonezije nevidna ločnica. Zahodno od nje je našel »azijske« živali, vzhodno pa »avstralske«. Ta ločnica, ki ji danes pravimo Wallaceova linija, je postala ena prvih opisanih meja med globalnimi biogeografskimi regijami, Wallace pa danes velja za očeta biogeografije. Med popotovanji po Južni Ameriki in Aziji se je Wallaceu – sočasno s Charlesom Darwinom – porodila tudi ideja o evoluciji z naravno selekcijo. Delo obeh naravoslovcev in njuno uspešno sodelovanje sta v sledečih desetletjih pripomogla k razvoju in uveljavitvi evolucijske teorije.
Razumevanje biogeografskih vzorcev in procesov, ki vplivajo na geografsko razširjenost vrst, se je od Wallaceovega časa do danes korenito spremenilo. Biološka disperzija (tj. razširjanje po prostoru) je predmet mnogih znanstvenih študij najrazličnejših organizmov, od palm do dinozavrov. Raziskave pogosto vključujejo tudi rekonstrukcijo preteklih disperzijskih poti z matematičnim modeliranjem. Kljub nedavnim teoretičnim in analitičnim napredkom pa imajo tovrstne rekonstrukcije pomembno metodološko pomanjkljivost: kako oceniti verjetnost disperzije na celostno informiran in biološko pomenljiv način?
Kaj je disperzija?
Pasivno ali aktivno premikanje po prostoru je značilno za tako rekoč vsa živa bitja, zato je presenetljivo, da je s tem povezana terminologija pogosto nejasna ter odvisna od vira in smisla uporabe. V ožjem smislu izraz disperzija pomeni hoten ali nehoten premik organizma, ki se ne ponavlja. Zanjo sta običajno značilni visoka stopnja smrtnosti in nizka stopnja nadzora smeri ter razdalje premika. Nameren, ponavljajoč, pogosto sezonski premik pa imenujemo migracija. Za razliko od disperzije sta za migracije značilni nizka stopnja smrtnosti in visoka stopnja nadzora poti ter cilja premika.
Poznamo različne tipe disperzije. Prva asociacija ob tej besedi je običajno tip, imenovan kolonizacija. Kolonizacija je redek ali celo enkraten premik med dvema območjema z ugodnim habitatom, med katerima je habitat neugoden (angl. long-distance dispersal). Znan primer kolonizacije je uspešno prečkanje 1000 kilometrov dolge morske poti med celinsko Južno Ameriko in Galapaškimi otoki pri prednikih današnjih galapaških želv (Chelonoidis niger). Najbližje živeče sorodnice galapaških želv (in najverjetneje tudi njihov skupni prednik) so kopenske želve, ki so slabe plavalke. Če takšno želvo morje slučajno odplavi z obale, je verjetnost njenega preživetja nizka – razen če jo, spet po naključju, naplavi nazaj na kopno. Vsaj eni breji samici (ali paru samca in samice) je v poznem oligocenu uspelo prav to: »po nesreči« preživeti pot na Galapaške otoke in tam uspešno ustanoviti novo populacijo – jih kolonizirati. Tak scenarij podpira tudi njihova biološka zasnova, saj so želve plovne in lahko več mesecev preživijo brez hrane in vode, poleg tega pa od obal Južne Amerike proti Galapaškim otokom teče Humboldtov morski tok.
Tovrstni izredni, naključni premiki med območji ugodnega habitata oziroma habitatnimi otoki vodijo v poseben tip biogeografskih vzorcev, ki mu s skupnim imenom rečemo otoška biogeografija. Zelo pomemben del otoške biogeografije je t. i. učinek ustanovitelja (angl. founder effect), močno zmanjšanje genetske raznolikosti populacije, ko ta izhaja iz majhnega števila prvotnih kolonizatorjev. Zaradi učinka ustanovitelja se lahko nova populacija genotipsko in fenotipsko zelo razlikuje od izvorne populacije, kar v ekstremnih primerih vodi v speciacijo, torej evolucijo nove vrste. Zaradi prostorske izolacije posameznih populacij galapaških želv danes poznamo dvanajst podvrst, mnoge od njih pa najdemo na le enem od otokov.
Kolonizaciji nasprotni proces je difuzija. Pri disperziji z difuzijo osebki postopoma napredujejo z robov areala populacije po ugodnem habitatu. Ta proces je zlasti nazoren (in pospešen) pri invazivnih vrstah. Primer je trsna krastača (Rhinella marina), ki je bila v tridesetih letih prejšnjega stoletja z namenom zatiranja škodljivcev na poljih sladkornega trsa s Havajev (kjer prav tako ni avtohtona) prinesena v Avstralijo. V naslednjih desetletjih je vrsta bistveno povečala svoj areal na severovzhodu Avstralije, z izpodrivanjem avtohtonih vrst pa je pustila močan pečat na naravni biološki raznovrstnosti območja.
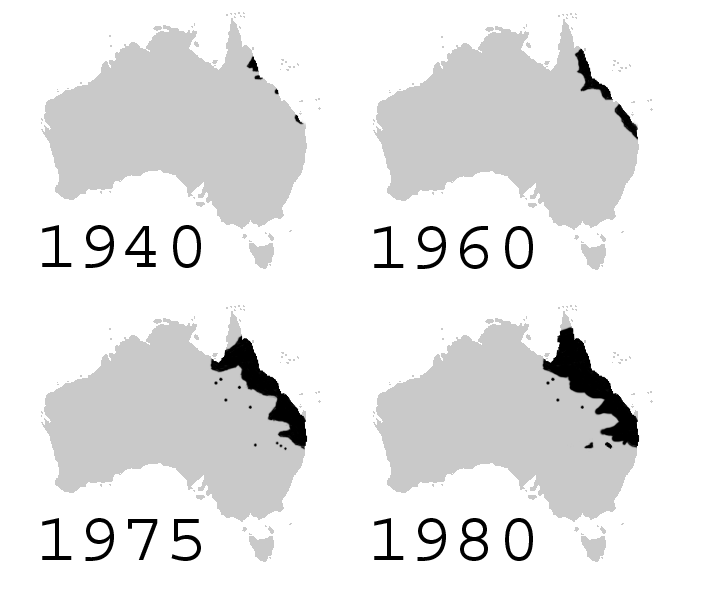
Zemljevid hitrega povečevanja njenega areala v Avstraliji (vir: Froggydarb, B kimmel, Wikimedia commons)
Disperzijo lahko klasificiramo tudi glede na vir energije, potrebne za premik. Aktivna disperzija je disperzija na »lasten pogon«, torej z organizmu lastnimi mehanizmi premikanja. Najjasnejši primer so aktivni letalci – ptice in netopirji naseljujejo skoraj vse kopenske habitate na svetu in so pogosto prvi kolonizatorji novonastalih otokov. Pasivna disperzija pa je na drugi strani značilna predvsem za nepremične organizme, kot so rastline. Pri pasivni disperziji organizem za premikanje izkorišča energijo iz okolja, pogosto veter ali morske tokove. Smer in razdalja premika sta tako nenadzorovana. Posebno zanimiv tip pasivne disperzije najdemo pri pajkih (na naslovni fotografiji prispevka). Balonanje (angl. ballooning) je poseben tip pajčjega vedenja, pri katerem se običajno mladi, lahki pajki na vetrovnih mestih postavljajo na konice nog, v veter spuščajo svilene niti in počakajo, da jih veter odpihne drugam. Večina takih poletov se konča relativno blizu, morda samo na sosednjem drevesu – včasih pa se zgodi, da pajki z vetrom prečkajo tudi oceane.

Pasivna disperzija z vetrom pri regratu (foto: Sandra Seitamaa, Unsplash)
Evolucijska prednost disperzije je fizični odmik od drugih, zlasti genetsko podobnih osebkov, kar zmanjšuje verjetnost parjenja v sorodstvu in tekmovanja za vire. Po drugi strani je cena disperzije tveganje za smrt med potjo, možnost neugodnih življenjskih pogojev na cilju ali trk ob disperzijsko pregrado. Te so lahko najrazličnejših oblik, na primer morje, reka ali gorska pregrada, kar je odvisno od biologije posameznega organizma. Na splošno je disperzija uspešna, kadar osebek uporabi »primerno« metodo transporta, preživi pot in v novem habitatu ustanovi populacijo, sposobno preživetja. Če novonastala in matična populacija ne vzdržujeta genetskega pretoka, sčasoma pride do speciacije.
Rekonstruiranje biogeografske zgodovine organizmov
Geografsko razširjenost vrst preučuje biogeografija. Ekološka biogeografija se ukvarja predvsem z današnjo razširjenostjo organizmov, zgodovinska biogeografija pa z njihovim geografskim izvorom in disperzijo skozi evolucijsko zgodovino. O zgodovinski distribuciji organizma lahko sklepamo le iz posrednih dokazov, kot so današnja distribucija sorodnih vrst, vzorci genetske strukture osebkov iz različnih populacij in (če je na voljo) fosilni material. Matematično modeliranje biogeografske zgodovine tarčnih organizmov (v nadaljevanju taksonov) v grobem upošteva tri vrste podatkov.
a) Prvi podatek je časovno kalibrirana filogenija. Filogenija je hipoteza o evolucijski zgodovini vključenih taksonov in služi kot ogrodje za rekonstrukcijo biogeografske zgodovine. Pri časovno kalibriranih filogenijah oziroma kronogramih je dolžina vej filogenetskega drevesa sorazmerna s pretečenim časom. Čim natančnejša časovna kalibracija, idealno podprta s fosili znanih starosti, je ključna za zanesljivost biogeografskih rekonstrukcij.
b) Drugi podatek je današnja razširjenost vključenih taksonov. Vsakemu taksonu je potrebno pripisati eno ali več biogeografskih regij, ki jih naseljuje danes. Izbira in razdelitev celokupnega areala vseh taksonov v biogeografske regije sta odvisni od specifične biologije taksonov in raziskovalnega vprašanja.
c) Tretji upoštevani podatek pa so verjetnosti disperzije med biogeografskimi regijami, določenimi v prejšnjem koraku. Ta tip vhodnih podatkov je pri biogeografskih rekonstrukcijah najmanj formaliziran in pogosto spregledan. Problematiko in pomembnost premišljenega ocenjevanja verjetnosti disperzije smo obravnavali v več študijah pajkov iz družine zlatih mrežarjev, predstavljenih v nadaljevanju (Turk idr. 2020; Turk idr. 2021; Kuntner in Turk 2022).
Osvetlitev problema ocenjevanja verjetnosti disperzije na primeru zlatih mrežarjev
Zlati mrežarji (Nephilidae) so družina tropskih in subtropskih pajkov, znani po nenavadnih fenotipih. Med njimi najbolj izstopa ekstremni velikostni spolni dimorfizem, tj. izjemna razlika med telesno velikostjo samcev in samic. Pri eni od vrst, orjaškem mrežarju (Nephila pilipes), so samice do petstokrat težje od samcev, kar je eden najekstremnejših primerov velikostnega spolnega dimorfizma pri kopenskih živalih. Zelo velike so lahko tudi asimetrične mreže, ki jih pletejo samice, kar imenujemo gigantizem mrež. Zlati mrežarji so biogeografsko zanimivi zaradi domnevno zelo davnega izvora (v mezozoiku pred 133 milijoni let), široke današnje razširjenosti na petih celinah in raznolike disperzijske biologije.

Majhen samec orjaškega mrežarja (Nephila pilipes) na hrbtu bistveno večje samice in primer gigantske mreže samice zlatih mrežarjev (foto: Matjaž Kuntner)
S problematiko ocenjevanja verjetnosti disperzije med biogeografskimi regijami smo se prvič srečali pri študiji, objavljeni leta 2020 v reviji Journal of Biogeography. Ugotavljali smo biogeografsko zgodovino celotne družine zlatih mrežarjev, torej njihov geografski izvor in potek razširjanja areala v zadnjih 130 milijonih let. Na voljo smo imeli novo, dobro podprto in časovno kalibrirano vrstno filogenijo, ki smo jo za čim boljšo taksonomsko reprezentacijo še dodatno razširili s tremi prej nevključenimi vrstami. Današnja razširjenost družine je dobro dokumentirana. Vsaki vrsti smo pripisali eno ali več biogeografskih regij, ki jih naseljuje danes (na spodnji sliki): neoarktično, neotropsko, paleoarktično, afrotropsko, indomalajsko in avstralazijsko ter otoke zahodnega Indijskega oceana, ki smo jih zaradi visoke stopnje endemizma zlatih mrežarjev na tem območju privzeli kot samostojno enoto.
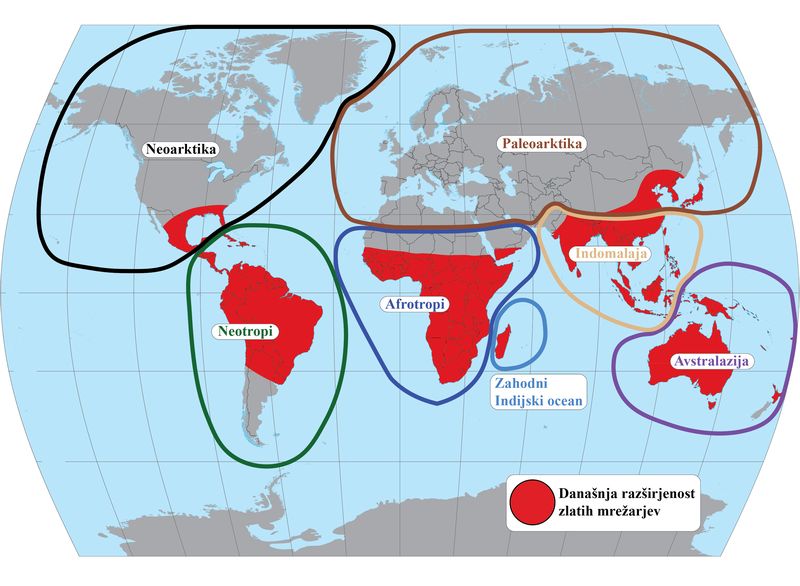
Sedem biogeografskih regij – današnja distribucija zlatih mrežarjev je prikazana v rdeči barvi (povzeto po Turk idr. 2020)
Zadnji vhodni podatek rekonstrukcijske analize, ki je sicer neobvezen, je matrika verjetnosti disperzije med biogeografskimi regijami. Verjetnosti so lahko poljubne vrednosti na skali od 0 do 1. V programu RASP, ki ga uporablja večina biogeografskih študij, je analizo mogoče tudi časovno razslojiti in za vsak par regij določiti lastno verjetnost disperzije za posamezni časovni sloj. Pri starejših študijah je bilo določanje verjetnosti disperzije običajno arbitrarno in grobo, predvsem pa subjektivno. Glede na velik potencialni vpliv disperzijske matrike na rezultat rekonstrukcije smo svoje ocene verjetnosti želeli osnovati na konkretnih, merjenih podatkih. Kot najprimernejšo osnovo za izračun ocen verjetnosti pri naši študiji smo izbrali najkrajšo fizično razdaljo med vsakim parom regij v posameznem časovnem sloju. Na računalniškem rekonstrukcijskem modelu premikanja tektonskih plošč GPlates smo izmerili razdaljo med najbližjima točkama vsakega para regij na vsakih 10 milijonov let, za časovno obdobje med 130 in nič milijoni let nazaj. Končnih 294 meritev smo pretvorili v verjetnost disperzije. Če sta bili regiji v določenem časovnem sloju v fizičnem stiku, smo določili najvišjo, 95‑odstotno verjetnost disperzije, kar pomeni, da je ta zelo verjetna, a ne nujna. Najnižjo, 5-odstotno verjetnost smo določili za razdalje nad 4000 kilometrov. To je približna razdalja med najodročnejšimi kraji, naseljenimi s pajki, in njim najbližjo celino (npr. med Havaji in Severno Ameriko). Med tema ekstremoma smo predpostavili padajočo linearno zvezo med razdaljo in verjetnostjo disperzije. Statistično najbolje podprti biogeografski model je izvor zlatih mrežarjev umestil v indomalajsko in avstralazijsko biogeografsko regijo.
S tem smo kot prva nam poznana raziskovalna skupina naredili resnejši korak k vpeljavi informiranih ocen verjetnosti disperzije v biogeografske rekonstrukcije. A verjetnost disperzije seveda ni odvisna le od bližine sosednjega ugodnega habitata, temveč tudi od številnih drugih biotskih in abiotskih faktorjev, ki jih ni vedno enostavno kvantificirati. Pri naslednji študiji smo želeli iti korak dlje in v ocene vključiti še več relevantnih informacij. Tokrat smo se osredotočili na en sam rod zlatih mrežarjev, rod Herennia. To je vrstno najbolj pester rod zlatih mrežarjev, ki je razširjen v tropski Aziji in Avstralaziji. Za večino od 14 poznanih vrst je značilen majhen areal, ki je pogosto omejen na en sam otok ali skupino otokov. Le ena vrsta, H. multipuncta, ima zelo široko razširjenost in je sinantropna, tj. živi blizu ljudi in človeških bivališč, medtem ko druge vrste živijo v neokrnjenih gozdovih. Razlog za dramatično razliko v velikosti arealov med to in drugimi vrstami ni znan, poleg tega ostaja nedokumentiran tudi glavni način njihove disperzije. Geološka zgodovina areala rodu je zelo kompleksna in zato zahtevna za natančno rekonstrukcijo.
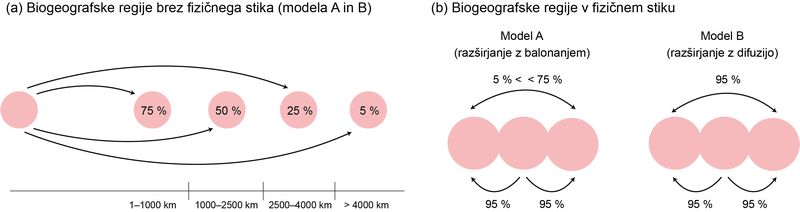
Metoda atribucije verjetnosti disperzije pri parih biogeografskih regij brez fizičnega stika (a) in v fizičnem stiku (b) (povzeto po Turk idr. 2021)
Ponovno smo imeli na voljo dobro podprto vrstno filogenijo in dobro poznavanje današnjega areala rodu. Verjetnosti disperzije med regijami, ki v določenem časovnem sloju niso bile v fizičnem stiku, smo ocenjevali enako kot prej, torej na podlagi minimalne fizične oddaljenosti. Poleg rekonstrukcijskega modela GPlates smo ocene verjetnosti informirali tudi s podatki iz geološke literature, predvsem o časovnih okvirih kopenskih mostov med različnimi otoki in celino. Zaradi kompleksnosti geološke zgodovine območja in posledične nezanesljivosti geoloških modelov smo verjetnosti disperzije med nepovezanimi regijami omejili na le štiri kategorične vrednosti. Fizični stik med regijami pa je bil obravnavan na dva različna načina. Model A je 95-odstotno verjetnost disperzije pripisal le parom regij v neposrednem stiku, model B pa je tolikšno verjetnost pripisal vsem regijam, ki so med seboj fizično povezane, torej tudi posredno povezanim parom regij. S tem smo poskušali oceno verjetnosti disperzije vsaj delno informirati tudi z biologijo organizma. Model A je predpostavljal kolonizacijo z balonanjem kot primarnim načinom disperzije, model B pa z difuzijo.
Rekonstruirana zgodovinska razširjenost se med biogeografskima modeloma A in B v večjem delu filogenije ni razlikovala. Obe analizi sta ugotovili široko izvorno razširjenost rodu v Avstraliji, celinski jugovzhodni Aziji in na Filipinih, nato pa postopno kolonizacijo preostalih regij.
Biogeografija naslednje generacije
S soavtorji smo o problematiki ocenjevanja verjetnosti disperzije v biogeografskih rekonstrukcijah razmišljali naprej – v nedavni objavi smo izluščili štiri glavne tipe informacij, ki bi jih biogeografija naslednje generacije, kot jo imenujemo, morala upoštevati za kar se da zanesljivo biogeografsko rekonstrukcijo: geološko zgodovino, geografske dejavnike, ekologijo in disperzijske lastnosti posameznega organizma. Pri tem je hkrati pomembno, da metodologija ohranja svojo fleksibilnost in tako omogoča uporabo na različnih organizmih z lastnimi specifikami. V prihodnjih letih bomo zato nadaljevali z razvijanjem biogeografske metodologije in oblikovali konkretne protokole za izračun najbolj pomenljivih ocen verjetnosti disperzije do sedaj.